содержание .. 1 2 3 4 5 6 7 8 9 10 ..
с. Механизм образования и принцип построения
химерных форм
Теперь уместно перейти к объяснению химер и, соблюдая историческую
последовательность, начать с работ Баура (Baur, 1910а, 191 Oh).
При этом необходимо отметить, что предлагаемый ниже
разбор отнюдь не является просто сводкой, но это необходимый фактический
материал для понимания той дискуссии, которая велась и обязательно будет
еще вестись в связи с установлением взаимоотношений между компонентами
химеры, с механикой их развития и даже с основными конструкциями химер.
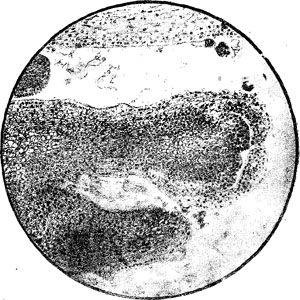
Рис. 14. Трихламидная химера тех же компонентов, с тем же относительным их расположением. В точке роста побега и с боков стебля видны три крупноклетных слоя с более темно окрашенными ядрами Баур оперировал с Pelargonium zonale, имеющей в естественном своем виде листья с белой каймой. Баур наблюдалг что у этой пеларгонии иногда появляются на кусте особые-ветки, несущие или чисто зеленые, или чисто белые листья (Кренке, 1928, стр. 451). Первые простым черенкованием, а вторые — прививкой могут быть легко выращены в соответствённые чип то зеленые или чисто белые кусты. Отсюда вытекала мысль, что Pelargonium zonale представляет собою естественную химеру. Исследовав конус нарастания этой пеларгонии, Баур нашел, что он состоит из разных тканей, именно из ткани с белыми пластидами и из ткани с зелеными пластидами.

Рис. 15. Деталь рисунка 14 в увеличенном виде. Видно, что верхние части заложений листьев не содержат в себе клеток внутреннего компонента.
Взаимное расположение этих тканей может быть
различно: либо один или два слоя белопластидных клеток облегают снаружи
зеленую часть конуса, либо наоборот. Кроме того, преимущественно
искусственным путем можно получить ,-конус нарастания «полузелёный
полубелый», с секториальным
распределением белой и зеленой ткани. Из конуса нарастания развиваются
побеги, листья и, наконец, цветы с половыми элементами. Если конус имел
периферические слои, состоящие из белопластидных клеток, то листья
развившегося из него побега также оказываются из двух компонентов
тканей: зеленая внутренняя часть облачена в белое покрывало (о числе его
слоев — ниже).

Рис. 16. Другой индивидуум трихламидной химеры.
Прекрасно видны три крупноклетных слоя как в точке роста побега, так и с
боков последней.

Рис. 17. Третий индивидуум трихламиДиой химеры Solanum iycopersi-cum — Solanum memphiticum.
При обратном отношении тканей то же получается и в
листе. Наконец, при секториально-разном конусе нарастания получаются и
растения с оригинальными секто-риально-пестрыми бело-зелеными листьями.
Первые два типа химер были названы периклинальными, а третий —секториадьным.
Периклинальные, в свою очередь, были разделены (Meyer, 1915) на: 1)
гаплохламидные, у которых один из компонентов облачен в ткань другого
компонента, состоящую всего из одного слоя клеток, и 2) диплохламидные,
у которых наружный компонент конуса нарастания состоит из двух слоев
клеток. С этим важным разделением, которое, по существу, было замечено и
Бауром, мы встретимся дальше, а здесь отметим новый, нами установленный
тип химер.
Среди наших химер мы впервые обнаружили трипдохламидные химеры, т. е.
такие, у которых конусы нарастания глав ных побегов при их заложении
покрыты тремя слоями клеток внешнего партнера (Solanum memphiticum Mart),
а срединная часть принадлежит внутреннему партнеру (Solanum lycopersicum
L.).

Рис. 18. Конусы нарастания главного и пазушного побегов прививочной трлхламидиой химеры, где Solanum memphiticum (Mart) тремя ясно видимыми слоями своих клеток облегает Solanum lycopersicum L.
Ввиду новизны этого сообщения мы приводим серию микрофотографически х
иллюстраций, как триплохламидных, так, для сравнения, и гаплохламидных и
диплохламидных (рис. 9—18 с объяснениями к ним). Триплохламидные химеры
резко отличались и в общей их морфологии и в интенсивности развития
(рис. 19, 1, и 20, II). Несколько подробнее мы остановимся на этом ниже.

Рис. 19. Solanum lycopersicum — Solanum memphiticum.
1—трихламидная химера. II — дихламидная химера. Привиты (
->) на Solanum memphiticum. Возраст химер и время прививок одинаковы, описание листьев дано в тексте.
Надо сказать, что мы не припоминаем углубленных рассуждений на тему,
почему именно из периклинальных химер до сих пор обнаружипилис.ь только
гашхо- и диплохламидные химеры. Наиболее носким основанием к этому могло
бы служить показание о соответствующей механике развития роста стебля
«двудольных» растений. Однако когда мы обратились к классической
литературе по этому вопросу (Hanstein, 1868, 1870; а задним Schoute,
1903; Kniep, 1904; Flot, 1906, 1907; затем Sachs, 1878; Errera, 1886;
Berthold, 1886; Haberlandt, 1891, и др.), то не нашли убедительных
данных, которые мешали бы принять возможность образования три- и более
хламидных химер. Более того, нередко трудно разграничить периблему от
плеромы, следовательно не в этой границе суть дела.

Рис. 20. Другие индивидуумы тех же химер (т. е.
Solanum lycopersicum — Solanum memphiticum), что и на рис. 19.
I — дихламидная химера (укорененный черенок) .II — трихламидная химера
привита на Solanum memphiticum.
Только дерматоген, как правило, остается резко ограниченным, но, как
увидим ниже, и здесь есть исключения. Наши, правда (Кренке, 1928, стр.
194—199; Krenke, 1933, SS. 236— 239) предварительные, исследования
заложения конусов нарастания побегов в каллюсе также не дали никаких
оснований, которые препятствовали бы признанию образования
полихла-мидных химер. Что же касается заложения химерных точек роста в
каллюсах, то здесь уже вовсе нет никаких оснований искать препятствий к
образованию полихламидных химер. Вместе с тем Кахидзе (1932) посредством
рентгенизации получила только гаплохламидные химеры, и она подчеркивает
это. Об этой работе будет сообщено в главе о стимуляционных химерах.

Рис. 21. Дихламидная химера Solanum lycopersicum L. — Solanum
memphiticum (Ц).
1 — побег гаплохламидной химеры (томат внутри).
2 — побег Solanum memphiticum. ch — химерная часть
двойного стебля (автосращение параллельного роста — см. 19-23 стр. 87, а
не мериклинальная структура). Продолжаясь далее самостоятельно, эта
часть выявляет периклинальнуго химеру S. L — чисто томатная часть
двойного стебля. Точка роста этой части заглохла на уровне надписи, t —
последний листок чистого томата. b, г — листья
химеры. Их гофрированность и чашевидность (например d, е) вызваны
несогласованностью роста обоих компонентов. Т — подвой — Solanum ,
lycopersicum. <---место прививки — Solanum memphiticum.