содержание .. 1 2 3 4 5 6 7 8 9 10 ..
МОРФОЛОГИЯ КЛЕТОК ДРОЖЖЕЙ
Размножение, рост и развитие культур дрожжей.
Одним из последствий
морфолого-физиологических изменений, объединяемых понятием развитие,
является размножение — воспроизведение новой клетки, тождественной с
материнской.
У большинства одноклеточных организмов следствием размножения является
увеличение числа организмов в популяции. Процесс размножения клеток
микроорганизмов обусловлен прежде всего «взаимосвязью» их с питательной
средой, в которой создаются благоприятные физико-химические условия,
регулирующие действие соответствующих ферментов. Установлено, что в период
задержки роста клеток дрожжей выделяемые в среду мощные восстановители
(редуцирующие ферменты), действуя на дисульфидные связи белка клеточной
оболочки, делают ее проницаемой для питательных веществ из среды, чем и
обеспечивается индуцированный биосинтез ферментов, в том числе
конститутивных [77].
Размножение. В зависимости от условий культивирования клетки дрожжей
размножаются вегетативным и' половым способами. Форма вегетативного способа
размножения определяется принадлежностью к той или иной биологически
самостоятельной группе — семейству — почкованием, делением, посредством
почкования, завершающегося делением.
В результате цитологического изучения функциональной морфологии клеточных
оболочек дрожжей, электронно-микроскопического исследования «родовых
шрамов», или рубцов, остающихся на месте отделения дочерней клетки от
материнской, описано 4 типа вегетативного размножения клеток дрожжей (рис.
18) [110]:
первый тип — у почкующихся дрожжей, например, семейства Saccharomycetaceae,
рода Saccharomyces, когда материал оболочек материнских клеток
непосредственно не переходит на обо-

лочки дочерних клеток. Клетки в начальных стадиях своего роста получают тот
материал клеточных оболочек, который синтезируется и накапливается у
основания образующейся почки.
Рубцы располагаются в областях максимальной кривизны клеточной оболочки,
часто тесно примыкая один к другому, по кругу или по спирали;
второй тип — у дрожжей семейства Saccharoinycoaaceae родов Saccharo-mycodes
и Hanseniaspora, особенностью которых является полярное почкование с
образованием множественного рубца. Почка образуется путем выпячивания
почечного рубца, оставшегося от предыдущего почкования, и растяжения
межклеточной перегородки;
третий тип — у делящихся дрожжей семейства Schizosaccharomyceta-сеае,
например рода Schizosaccharomyces, для которых характерным является
биполярный рост клеточных оболочек. Новый материал стенок откладывается от
центра к концам клетки в виде кольцеобразного выроста в месте ее деления;
четвертый тип — принадлежит цилиндрическим клеткам Endomyces magnusii.
Структурные изменения при делении происходят на всей оболочке цилиндрической
клетки. Если делений много, то оболочка состоит из нескольких пластин,
расположенных параллельно и заключенных одна в другой на рубце.
В процессе вегетативного размножения дрожжей из материнской клетки в
дочернюю переходит часть структурных образований цитоплазмы и ядра. При этом
перед каждым клеточным делением в ядре хромосомы воспроизводятся все
одновременно и по одному разу, чем и обеспечивается равное распределение их
между материнской и дочерней клеткой. В основе превращения хромосом лежит
молекулярный механизм воссоздания ДНК-При таком типе деления ядра,
называемом митозом, в поколениях клеток сохраняется строго постоянное число
хромосом [63].
Если рассматривать размножение дрожжей во времени, то в оптимальных условиях
примерно за 1 ч происходит полное формирование новой дочерней клетки. Однако
одна дрожжевая клетка не может бесконечно повторять процесс почкования.
На протяжении всего периода жизни материнская клетка среднем имеет 25—30
родовых шрамов, т. е. почкований. Сдерживающим фактором является изменение
структуры клеточной оболочки, связанное с образованием рубцов, уменьшающих
полезную поверхность материнской клетки, что ведет к снижению обмена веществ
и содержания РНК, ДНК, протеина в клетках и в конечном счете к гибели.
Половой способ размножения дрожжей, связанный с прорастанием аскоспор (спор,
находящихся в сумках или асках) в вегетативные клетки, сопровождается
мейотическим процессом деления ядра.
Резкий переход культивирования дрожжей из полноценной среды в среду, бедную
питательными веществами, при достаточной влажности, значительном накоплении
запасных веществ в клетке и доступе кислорода воздуха приводит к
спорообразованию. Аскоспоры устойчивы к неблагоприятным внешним условиям,
действию высокой температуры, высушиванию, но они менее термостабильны, чем
бактериальные споры, и погибают при температуре 60°С, а споры бактерий
выдерживают температуру кипящей воды и более высокую.
Аскоспоры обычно образуются в результате предшествующего полового слияния
двух дрожжевых клеток и деления оплодотворенного ядра. В одном аске
образуются 1—4, иногда 8 спор. При условиях, благоприятных для вегетативного
развития, на свежей питательной среде споры прорастают и снова превращаются
в почкующиеся клетки.

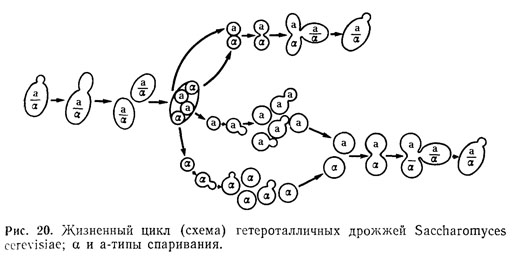
Жизненный цикл у дрожжей связан с чередованием вегетативного размножения и
спорообразования с разной продолжительностью гаплоидной и диплоидной стадий
(рис. 19). Дрожжи-сахаромицеты со сменой дипло- и гаплофаз делятся на 2
группы: гетероталличные и гомоталличные [64].
Гетероталличные штаммы дрожжей имеют устойчивые диплоидную и гаплоидную
фазы. Диплоидные клетки могут неограниченно долго размножаться вегетативно,
переходить при неблагоприятных условиях к спорообразованию, в результате
чего образуются аски с гаплоидными спорами, каждая из которых принадлежит к
одному из двух типов спаривания (а и а). При этом копуляция совершается лишь
при встрече двух аскоспор или клеток из разных гаплоидных штаммов,
происходит образование зигот и восстановление диплофазы (рис. 20) [63].
Природа различия клеток а и а обусловлена их способностью вырабатывать
соединения гормональной природы. Предполагается, что «половой гормон»
увеличивает активность ферментов, разрушающих клеточную стенку. Реакция
клеток, предшествующая их копуляции (удлинение клеток), может проявляться
при помещении клеток разных типов спаривания на некотором расстоянии одна от
другой [64].
Гомоталличные штаммы дрожжей отличаются от гетероталличных тем, что имеют
только устойчивую диплофазу. Гаплоидные споры, изолированные из аска,
образуют диплоидную культуру, т. е. в потомстве отдельной споры происходит
слияние гаплоидных клеток — самодиплоидизация, за счет слияния спор в любых
комбинациях сестринских гаплоидных клеток или материнской клетки со своей
почкой. Типы спаривания существуют и у гомоталличных дрожжей [64, 121]. А.
Ф. Руснак, проанализировав большой набор рас дрожжей вида Sacch. vini,
используемых в виноделии, сделала вывод о преимущественно гомоталличной
природе их [169].
Однако некоторые дрожжи, например Zygosaccharomyces и Schizosaccharomyces,
длительное время размножаются вегетативно, находясь в гаплоидном состоянии.
Перед спорообразованием гаплоидные клетки сливаются и образуется диплоидная
зигота, которая делится путем мейоза и дает начало 4 или 8 гаплоидным
спорам. Споры эти прорастают и начинают размножаться бесполым путем в
гаплоидном состоянии.
При спорообразовании замедлен обмен веществ и общая жизнедеятельность
микроорганизмов. Такое состояние обеспечивает их выживаемость в условиях,
неблагоприятных для вегетативного размножения. Поэтому спорообразование,
объединяющее в себе процесс размножения и сохранения вида, следует
рассматривать как положительную стадию в индивидуальном развитии дрожжевых
организмов [83].
Интересные результаты получены Б. Пазоньи [290] по влиянию половой стадии
размножения у дрожжей на их жизненность и продуктивность. Культуры винных
дрожжей, прошедшие массовую споруляцию с последующим прорастанием спор и
диплоиди-зацией, превосходят те же культуры, не прошедшие этих стадий, по
быстроте сбраживания виноградного сусла, и при испытании в
полупроизводственных условиях наблюдалось сокращение периода главного
брожения с 35 до 21—31 сут.
Рост и развитие культур дрожжей. Микробные культуры в жидкой питательной
среде развиваются по кривой. Наблюдение за ходом ее, сопровождаемое
определением морфо-лого-физиологических и химических изменений в клетках и
вереде, является основным методом микробиологии [65, 151]. Фазы роста на
определенных участках кривой могут свидетельствовать до некоторой степени об
определенных стадиях развития и физиологическом состоянии культуры дрожжей
(рис. 21). В общем виде кривая роста приведена на рис. 22. В периодических
условиях культивирования различают 4 основные фазы роста.
Лаг-фаза — это период с момента внесения посевного материала в питательную
среду до установившейся постоянной скорости роста культуры. Эта фаза
включает латентный (скрытый) этап, когда микроорганизмы приспосабливаются к
новым условиям и некоторое число клеток может даже погибнуть, и этап начала
роста — этот период характеризуется интенсивной метаболической активностью,
хотя число организмов в культуре или совсем не увеличивается или
увеличивается незначительно. Наблюдается заметное увеличение размеров
клеток, возрастает содержание в них общего белка, нуклеиновых кислот,
энергично синтезируются адаптивные ферменты. Чем полноценнее среда и моложе
культура, тем короче лаг-фаза. Если вносят значительное количество молодого
посевного материала, то культура развивается без лаг-фазы [151].

Логарифмическая, или экспоненциальная, фаза характеризуется максимальной
скоростью размножения дрожжей, самой быстрой для данных условий. В этот
период численность клеток и их суммарная биомасса возрастают в
геометрической прогрессии. Средний размер клеток дрожжей становится
минимальным, большинство из них — почкующиеся, с однородной цитоплазмой и
тонкой оболочкой. Для них характерна физиолого-биохимическая активность и в
то же время они более чувствительны к действию различных неблагоприятных
факторов, чем зрелые и покоящиеся.
Культура состоит из «стандартных клеток». Однако длительность
экспоненциальной фазы роста для периодических условий культивирования на
жидких питательных средах невелика, так как питательные вещества
потребляются из среды, а в ней накапливаются ненужные продукты обмена, т. е.
среда постепенно становится менее благоприятной для роста, в результате чего
культура переходит в стационарную фазу.
Чтобы стабилизировать культуру в одном и том же состоянии, применяют
проточные, непрерывно обновляемые среды.
В непрерывно обновляемой культуре устанавливается динамическое равновесие
между составом среды и количеством клеток. Метод непрерывных культур имеет
практическое применение в генетике: он позволяет наблюдать за появлением и
исчезновением мутантных форм в популяции; в бродильной и микробиологи ческой
промышленности он используется при производстве продукции определенного
состава.
Стационарной фазе свойственно постоянное число живых организмов и
максимальная плотность популяции. В этот период число погибающих клеток
становится равным числу вновь образующихся, устанавливается уравновешенное
размножение их и отмирание.
Фаза отмирания клеток характеризуется уменьшением числа жизнеспособных
клеток в культуре, преобладанием мертвых, часто автолизованных. При брожении
виноградного сусла эта фаза проявляется после полного сбраживания сахара и
при выдержке виноматериала на дрожжевом осадке. В этой фазе наступает
старость культуры, которая сопровождается резкой
функционально-морфологической перестройкой клеток. Они становятся мельче,
часто деформируются, протоплазма приобретает зернистый вид. Особенно вид
клеток меняется при долгом пребывании дрожжей в осадке вина при недостатке
или полном отсутствии кислорода воздуха. Количество протоплазмы у
автолизую-щихся клеток постепенно уменьшается в результате расщепления
белков под влиянием собственных протеолитических ферментов, клетки
становятся почти пустыми с небольшими капельками жира. При длительном
пребывании клеток дрожжей в осадке вина с доступом кислорода воздуха можно
видеть покоящиеся формы, которые долго остаются жизнеспособными главным
образом за счет усвоения органических кислот.
Каждая фаза соответствует определенным скоростям роста, для выражения
которых пользуются абсолютными и относительными показателями. Абсолютную
(валовую) скорость роста культуры (U) за определенный отрезок времени
вычисляют по формуле [65, 152]


Величина экономического коэффициента зависит от характера энергетических
процессов. При дыхании этот коэффициент относительно высок, при брожении он
ниже и меняется в зависимости от условий.
Скорость роста дрожжей и ее лимитирование разнообразными факторами влияют на
кинетические свойства культуры (популяции), на состав клеток и состав
продуктов обмена веществ [152].
Рост культуры дрожжей на виноградном сусле определяется главным образом
четырьмя факторами: концентрацией продуктов обмена (этанола), температурой,
концентрацией растворенного кислорода и концентрацией клеток дрожжей.
Поскольку этиловый спирт всегда присутствует в бражке, а брожение протекает
в анаэробных условиях (концентрация растворенного кислорода практически
равна нулю) обычно при постоянной температуре, влияние этилового спирта на
рост культуры имеет решающее значение и основным уравнением кинетики
процесса является уравнение [27]:
m = 0,19 — 0,019
* P, где Р — концентрация этилового спирта, % об.
Следует отметить, что в литературе еще мало сведений, которые позволили бы
выявить зависимость образования определенных продуктов брожения от той или
иной скорости роста культуры (популяции) и вида фактора, ограничивающего
рост. Изучение это осуществимо только при непрерывном методе
культивирования, математическое описание которого позволяет отойти от
эмпирического подбора оптимальных условий [19, 32, 59,123]